Klamath IFRMP Conceptual Models
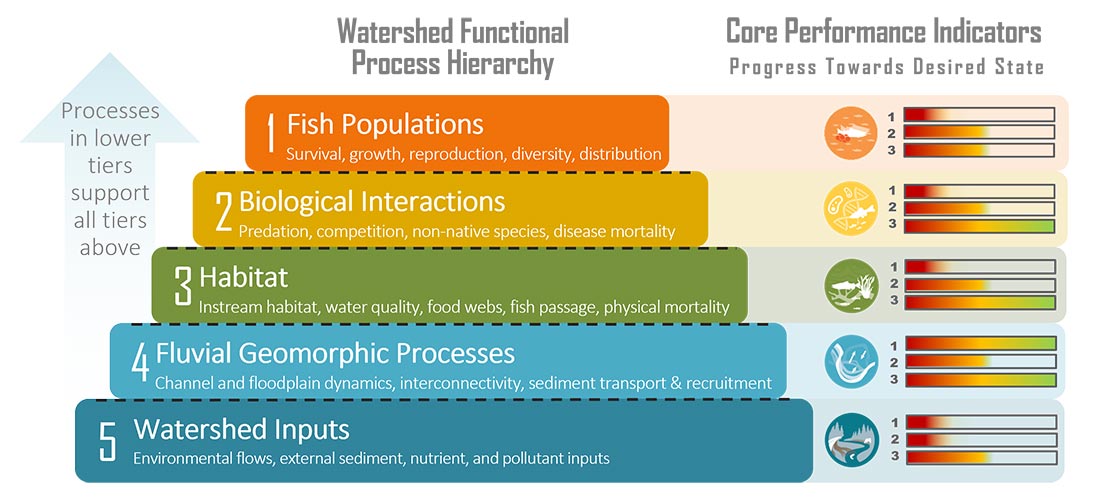
ESSA developed conceptual models for what we considered the primary drivers of habitat conditions for fish in the Klamath Basin (i.e. fluvial geomorphic processes and
This plan is meant to serve as a dynamic roadmap that describes the highest priority functional watershed restoration and monitoring actions that can help reverse the declines of multiple native Klamath Basin fish populations to help benefit ecosystems and communities.
These outputs were derived through a remarkably collaborative series of engagements by hundreds of basin participants 2016-2022.





ESSA developed conceptual models for what we considered the primary drivers of habitat conditions for fish in the Klamath Basin (i.e. fluvial geomorphic processes and
To assist with future monitoring and status reporting for the Klamath basin, ESSA researched potential suitability thresholds for candidate performance indicators and compiled available information
After careful consideration of alternatives, we adopted a multi-criteria scoring approach to prioritization that has undergone multiple rounds of peer-review by Sub-basin Working Group (SBWG)



The IFRMP (Plan) was completed in partnership with the Pacific States Marine Fisheries Commission and ESSA with extensive input from over 50 partners between 2016 and 2023. After seven years in the making and hundreds of hours of collaborative discussions with tribal, state, federal, and conservation partners, the IFRMP is ready for implementation. The Plan will serve as a dynamic roadmap describing the highest priority watershed restoration and monitoring actions to help reverse the declines of multiple native Klamath Basin fish populations to benefit ecosystems and communities. Focal fish species include Chinook and coho salmon, steelhead, bull trout, redband trout, Pacific lamprey, Lost River sucker (C’waam), shortnose sucker (Koptu), green sturgeon and eulachon.
“Impacts to native fish have been deeply felt by many who live, work, and fish across the Klamath Basin,” said Service Regional Director Paul Souza. “The IFRMP provides a unifying framework for coordinated restoration and recovery of native fish species from the headwaters to the Pacific Ocean, while improving flows, water quality, habitat and ecosystem processes for the benefit of local communities.”
The IFRMP is made up of several components including:
This work has delivered the vision of the Klamath Basin IFRMP to provide a unifying framework for planning coordinated recovery of native fish species from the headwaters to the Pacific Ocean while improving flows, water quality, habitat, and ecosystem processes. It is very rare to achieve the degree of sustained collaboration afforded by the IFRMP planning process and to emerge with a Basin-wide package of practical restoration and monitoring priorities.
The Plan is now available for use by all partners and stakeholders engaging in fisheries restoration and is intended to be updated periodically as restoration work is completed and as new ideas and information are brought forward.
All are to be commended for their efforts and the legacy of collaboration that was created.